As published in Human Reproduction Update 1996,
Vol.2, No. 4 pp. 295-306
Elizabeth A. Stewart, M.D. and Romana A. Nowak,
Ph.D.
Department of Obstetrics, Gynecology and Reproductive Biology, Harvard Medical School, Brigham and Women's Hospital, Boston, Massachusetts, USA
Leiomyomas are an important cause of menorrhagia and other forms of abnormal uterine bleeding. The pathogenesis of this process is largely unknown, however. A classic theory, first suggested by Sampson's work in 1912 (Surg. Gynecol. Obstet., 14, 215-230), states that local dysregulation of the vascular structures in the uterus is responsible for this abnormal bleeding. Recent work demonstrates dysregulation of a number of growth factors in the myomatous uterus. As many of these factors regulate the process of angiogenesis or have other effects on vascular structures, we hypothesize that this dysregulation of growth factors or their receptors provides the molecular mechanism underlying these vascular abnormalities. In turn, these abnormal vessels lead women with leiomyomas to experience menorrhagia. Factors that may prove to be important in this process include basic fibroblast growth factor, vascular endothelial growth factor, heparin-binding epidermal growth factor, platelet derived growth factor, transforming growth factor-b, parathyroid hormone-related protein and prolactin. Current treatment regimens for women with leiomyoma-related bleeding depend on the manipulation of the steroid hormone environment. By better understanding the pathogenesis of this disease process, therapies directed against growth factor abnormalities may result in better treatment with less harmful side-effects.
Key words: angiogenesis/basic fibroblast
growth factor/ leiomyoma/menorrhagia/uterus
Introduction
Leiomyoma-related menorrhagia is a significant medical and social problem for many women. Effective treatment strategies are limited by a narrow understanding of the pathogenesis of this disease. Classic studies suggest that there is a fundamental alteration in the vascular structures of the myomatous uterus. Recent progress in defining the molecular mechanisms of angiogenesis in the uterus lends support to this theory by demonstrating local dysregulation of vasoactive growth factors or growth factor receptors in leiomyomas or leiomyomatous myometrium. Thus, the molecular mechanisms underlying the process of leiomyoma-related menorrhagia are delineated.
Although ectasia of the venules is the best-characterized vascular abnormality in the myomatous uterus, multiple defects of arterioles, veins, and the extracellular matrix (ECM) surrounding them are probably responsible for this heterogeneous disorder. Understanding this dysregulation will not only define the pathophysiology of this important clinical problem, but it may also lead to innovative treatments.
This article will review what is known about the
process of normal menstruation and angiogenesis in the uterus as a basis for
understanding the molecular mechanisms underlying this process. After reviewing
the vasoactive growth factors that demonstrate altered expression in leiomyomas,
we will explore options for potential therapeutic modalities.
Normal menstrual function
Normal menstrual function is classically described by cyclic endometrial changes that result from the production of ovarian steroids. The basic histologic pattern that was first described in 1950 by Noyes, Hertig and Rock is still used today (Noyes et al., 1950). Both stromal and glandular components of the endometrium are affected by the cyclic changes in estrogen and progesterone during the normal menstrual cycle.
This progression of changes in the morphology of glands and stroma is parallelled by changes in the vascular structures. These changes start in the myometrium underlying the endometrium. Arcuate arteries arising in the myometrium give off radial branches (also known as basal arteries) that become coiled and form the so-called spiral arteries as they penetrate into the endometrium. The spiral arteries, unlike the basal arteries, are sensitive to estrogen and progesterone (Kaiserman-Abramof and Padykula, 1989). These vessels form the rich network of interdigitating capillaries that supply the functional layer of endometrium that is shed during menstruation.
Menstruation is preceded by an ischemic phase characterized by vasoconstriction of these spiral arteries and arterioles, and bleeding ensues after these vessels relax (Kaiserman-Abramof and Padykula, 1989). Blood components including clotting factors and platelets appear to form clots that limit blood loss until regeneration is complete.
The withdrawal of steroid hormones preceding
menstruation results in disruption of the endometrial cells and the
extracellular matrix (ECM) as well (Tabibzadeh, 1996). Disordered expression of
molecules including desmoplakin I/II, E-cadherin, a- and b-catenins
and loss of F-actin occurs only in the functionalis layer to allow for menstrual
shedding (Tabibzadeh et al., 1995). Apoptosis increases late in the
secretory phase in the endometrial glands and thus prepares this tissue for
disruption (Tabibzadeh, 1996). After the functional layer is shed, regeneration
proceeds from the endometrial basalis. With the basalis in direct contact with
the myometrium, a mechanism is established whereby myometrial growth factors
influence endometrial regeneration in a paracrine fashion.
Angiogenesis in the uterus
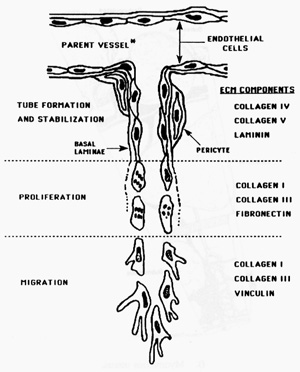
The constant cyclic process of angiogenesis, the formation of new blood vessels, in the ovary and uterus is unique and not well understood. Angiogenesis is usually seen in pathologic processes such as wound healing or tumour formation (Torry and Rongish, 1992), and the female reproductive tract provides a unique opportunity to study the physiology of this process. It involves an interaction between the blood vessels themselves and their surrounding ECM. The sequence of events involved in angiogenesis is (i) basement membrane degradation, (ii) endothelial cell migration, (iii) endothelial cell proliferation, and (iv) capillary tube formation and stabilization (Torry and Rongish, 1992; Figure 1).
Figure 1. A schematic diagram of the
events involved in angiogenesis: a capillary is diagrammed arising from a parent
vessel. The first step of basement membrane degradation is not depicted. Once
cells move through the basement membrane they migrate as a sprout in response to
an angiogenic stimulus. The ECM components, collagen IV and V and laminin are
associated with the basement membrane and entry into an area rich in the
interstitial collagens I and III and fibronectin encourages migration.
Proliferation follows migration by 24 hours. These angiogenic sprouts then
undergo vacuolization to form a capillary lumen. Once flow is established a new
basement membrane is formed around the capillary in the process of tube
stabilization. From Torry RJ and Rongish BJ. Angiogenesis in the uterus:
Potential regulation and relation to tumor angiogenesis. Am J Reprod Immunol
1992; 27:171-9. Reproduced with permission of the publisher, Munksgaard, Ltd.
Basement membrane degradation involves stromelysins,
collagenases, and other enzymes to degrade specific elements of the ECM (Torry
and Rongish, 1992). Endothelial cells can then migrate to the end of a vessel.
This migratory process is believed to be favored by an environment rich in
collagens type I and type III and to be stimulated by basic fibroblast growth
factor (bFGF; Torry and Rongish, 1992).
These ECM proteins also appear to be important in
the next step of proliferation (Torry and Rongish, 1992). Lumen formation and
stabilization are less well understood but also appear to be influenced by
components of the ECM.
Leiomyomas as a cause of abnormal uterine
bleeding
Leiomyomas (fibroids) are benign smooth-muscle tumors of the human uterus. They result from somatic mutations and clonal expansion of normal myometrial cells (Barbieri and Andersen, 1992). Leiomyomas are clinically important since they are a major source of abnormal uterine bleeding that leads to hysterectomy or other medical or surgical intervention. There is no well-established animal model for the study of these neoplasms. During 1988-1990, one-third of the approximately 1.7 million women who had hysterectomies each year in the United States had a diagnosis of uterine leiomyomas (Wilcox et al., 1994). Women of color are overrepresented in this group, with a primary diagnosis of leiomyoma in 61% of cases (Wilcox et al., 1994).
Approximately 30% of women with leiomyomas
experience menstrual abnormalities, with menorrhagia or excessively heavy menses
being most common (Buttram and Reiter, 1981). There is no evidence that this
abnormal bleeding is related to either an increased surface area of the
endometrial cavity or to increased incidence of ovulatory dysfunction (Buttram
and Reiter, 1981).
Abnormal vascular structure: Abnormal bleeding
The theory that best explains leiomyoma-related abnormal bleeding states that the primary event is a change in venous structures in the endometrium and myometrium resulting in venule ectasia (Farrer-Brown et al.,1971, Buttram and Reiter, 1981; Figure 2).
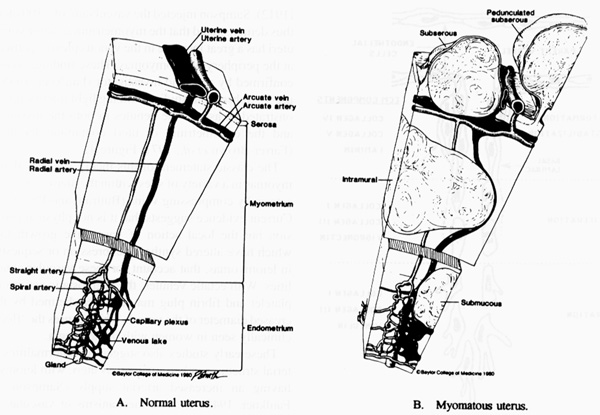
Figure 2. A diagrammatic representation of
a normal uterus and its vascular structure (A) and the venule ectasia found in
the endometrium and myometrium of the myomatous uterus (B) as proposed by Farrer-Brown.
This venule ectasia was ascribed to physical obstruction of the vessels by
leiomyomas. From Buttram VC, Reiter RC. Uterine leiomyomata: etiology,
symptomatology and management. Fertil Steril 1981: 36:433-45. Reproduced with
permission of the publisher, the American Society for Reproductive Medicine (The
American Fertility Society).
Among the evidence to support this theory of
vascular dysregulation in leiomyoma-related bleeding is the classic descriptive
study by Sampson (Sampson, 1912). Sampson injected the vasculature of over 100
uteri and thus demonstrated that the myometrium in leiomyomatous uteri has a
great increase in the venous plexus, particularly at the periphery of leiomyomas.
These findings were later confirmed by a second investigator (Faulkner, 1944;
Figure 3).
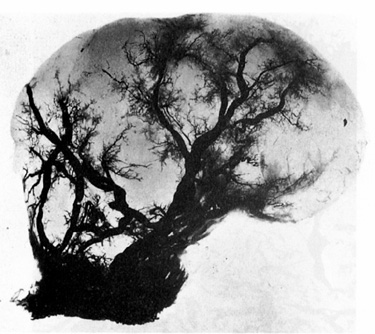
Figure 3. Photograph of a pedunculated
myoma where the venous system has been injected with neoprene. From Faulkner RL.
The blood vessels of the myomatous uterus. Am J Obstet Gynecol 1945; 47:185-197.
Reproduced with permission of the publisher, Mosby-Year Book, Inc.
Subsequent studies utilizing light microscopy
demonstrated ectasia of the venules in both the myometrium and the endometrium
of uteri containing leiomyomas. (Farrer-Brown et al., 1971; Figure 4).
The classic statement of this theory suggests
that leiomyomas in a variety of sites within the uterus cause venule ectasia by
compressing veins (Buttram and Reiter, 1981). Current evidence suggests that it
is not physical compression but the local action of vasoactive growth factors
which have altered synthesis, expression or sequestration in leiomyomas, that
account for these vascular abnormalities. With ectatic venules, the hemostatic
actions of the platelet and fibrin plug may be overwhelmed by the increased
diameter of the vessels which causes the "flooding" clinically seen in
women with menorrhagia.
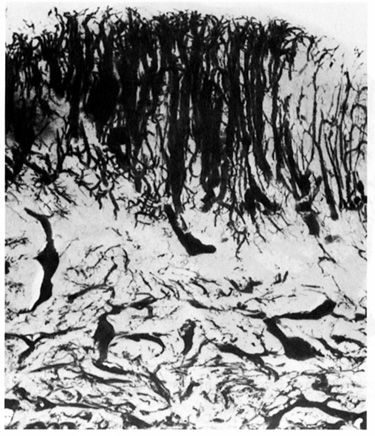
Figure 4. A microradiograph of a 300 m
cross section of endometrium from a myomatous uterus. Dilated veins are present
throughout the entire width of the endometrium. From Farrer-Brown G, Beilby JOW,
Tarbit MH. Venous changes in the endometrium of the myomatous uteri. Obstet
Gynecol 1971; 38:743-51. Reprinted with permission from the American College of
Obstetricians and Gynecologists.
These early studies also suggested abnormalities in
arterial structures in leiomyomatous uteri, with leiomyomas having an increased
arterial supply (Sampson, 1912; Faulkner, 1944). Several mechanisms of vascular
dysregulation are consistent with both the heterogeneous clinical presentation
of women with leiomyomas and the diversity of chromosomal abnormalities found in
this neoplasm (Rein et al., 1991). Thus, multiple somatic mutations can
result in the same phenotypic changes.
This paradigm of dysregulation of growth factor
function that results in disruption of normal vascular function appears relevant
for non-uterine bleeding disorders. Hereditary hemorrhagic telangiectasia is
characterized by profuse bleeding from mucosal surfaces including the
gastrointestinal tract and nasal epithelium, and by mucosal venule ectasia (Guttmacher
et al., 1995). The molecular defect causing this disease is a mutation in
endoglin, a binding protein that regulates the function of transforming growth
factor-b1 (TGF-b1; McAllister et al.,
1994).
Differential growth factor expression: myometrium
versus leiomyomas
The myometrium is a huge potential reservoir for paracrine or endocrine factors that regulate endometrial function. The apposition of these two tissues and the directional blood flow from the myometrium to the endometrium facilitate this interaction. Growth factors that stimulate angiogenesis or relax vascular tone and that have overexpression of either ligand or receptor in leiomyomatous uteri are candidates to cause abnormal uterine bleeding in these women and thus become targets for potential therapies. Alternatively, underexpression of angiogenic inhibitory factors or vasoconstricting factors or their receptors in leiomyomas may also result in abnormal bleeding.
It is clear that numerous genes are differentially regulated in neoplastic leiomyomas and in normal myometrium. Both the estrogen receptor and the progesterone receptor as well as the enzyme aromatase are constitutively overexpressed in leiomyomas compared with myometrium (Soules and McCarty, 1982; Rein et al.,1990; Brandon et al., 1993; Bulun et al., 1994). Leiomyomas also overexpress receptors for insulin-like growth factor-I (IGF-I) and mRNA for IGF-II (Tommola et al, 1989; Vollenhoven et al., 1993; Chandrasekhar et al., 1992 and Gloudemans et al., 1990) and have up to a sixfold increase in expression of TGF-b3 compared with matched myometrium (Murphy et al., 1994). Data from our laboratory also demonstrate the overexpression in leiomyomas of both mRNA and protein for parathyroid hormone-related protein (PTHrP) and bFGF (Weir et al., 1994; Mangrulkar et al., 1995).
Certain proteins in leiomyomas appear to undergo
menstrual cycle phase-specific upregulation compared with normal myometrium.
Work from our laboratory has demonstrated the relative overexpression of mRNAs
for collagen type I and collagen type III in leiomyomas only in the
proliferative phase of the cycle (Stewart et al.,1994). Epidermal growth
factor (EGF) mRNA has been shown to be relatively overexpressed in leiomyomas
during the luteal phase compared with matched myometrium (Harrison-Woolrych et
al., 1994). Earlier studies suggest the receptor for EGF may be decreased in
leiomyomas since binding studies show decreased binding in leiomyomas compared
with myometrium (Tommola et al., 1989).
Molecular mediators of leiomyoma-related bleeding
Growth factors or their receptors that are differentially regulated in the leiomyomas or the endometrium of leiomyomatous uteri are potential mediators of leiomyoma-related complications. These differentially regulated factors, which are known to act on vascular tissue by increasing proliferation or changing vessel caliber, are potential causes of leiomyoma-related menorrhagia. The factors that meet these criteria include bFGF, vascular endothelial growth factor (VEGF), heparin-binding epidermal growth factor (HBEGF), platelet derived growth factor (PDGF), TGF-b, PTHrP and prolactin.
Four of these factors (bFGF, VEGF, HBEGF, PDGF)
belong to the heparin-binding group of growth factors. Since these factors all
bind to heparin sulfate proteoglycans found in ECM, leiomyomas, with their large
ECM content may prove to be a reservoir for these factors. Both bFGF and VEGF
primarily regulate endothelial cell function and may thus promote the
endothelial cell migration vital to the angiogenic process. HBEGF and PDGF
primarily regulate fibroblast and smooth muscle cell function and may thus
influence vascular smooth muscle, leiomyoma or myometrial cells themselves, or
the endometrial stromal cells. PTHrP can act as a vasodilator either indirectly
through actions on the ECM or directly on vessels. TGF-b
acts on many cell types and prolactin, when cleaved, acts as an inhibitor of
angiogenesis. Thus, these factors represent a wide array of potential actions
that regulate vascular function in the uterus.
Basic fibroblast growth factor
Basic fibroblast growth factor (bFGF) is an 18-kd protein that promotes angiogenesis through a number of mechanisms, including the induction of endothelial cell proliferation, chemotaxis, and the production of matrix remodelling enzymes such as collagenase and plasminogen activator (Presta, 1988). In human endometrial transformed cell lines, estradiol treatment stimulates bFGF-like activity, which is lost when cells are treated by progesterone; this model mimics hormonal influences regulating angiogenesis in vivo (Presta,1988). bFGF has also been shown to be the major mitogen causing proliferation of vascular smooth muscle cells after injury (Linder and Reidy, 1991). bFGF has been detected by PCR in human endometrium throughout the menstrual cycle and shown by immunohistochemistry to localize primarily to the glandular epithelial cells (Ferriani et al., 1993).
It is known, however, that bFGF is stored in ECM bound to heparin sulfate proteoglycans, which may increase its local bioavailability (Vladovsky et al., 1987). Leiomyomas, which have large quantities of ECM, may serve as stores of heparin-binding growth factors which in turn may affect surrounding endometrial tissue function. Studies from our laboratory have shown that leiomyomas do indeed contain significant stores of bFGF (Mangrulkar et al., 1995). Immunohistochemistry for bFGF revealed only light staining within the myometrial, leiomyoma or vascular smooth muscle cells. Light staining was also present around the basement membrane. However, the large areas of ECM that distinguish leiomyomas from normal myometrium showed strong immunoreactivity for bFGF. Leiomyomas also displayed higher levels of bFGF mRNA than corresponding normal myometrium (Mangrulkar et al., 1995). Thus, leiomyomas synthesize and store significantly more bFGF than normal myometrium.
bFGF can act through several different receptors but most commonly acts through the type I receptor [encoded by the flg or fms-like gene (FGFR-1)] or the type II receptor [encoded by the bek gene (FGFR-2)] (Fernig and Gallagher, 1994). The two alternatively spliced isotypes of FGFR-1 have been localized to the human fetal uterus (Yeh and Osathanondh, 1993). Recently we reported that both these forms are present in adult uterine endometrium, myometrium and leiomyomas throughout the menstrual cycle (Anania, et al., 1995). However, immunohistochemical staining suggests however that there is differential expression of these receptors within the uterine tissues as the menstrual cycle progresses. Immunoreactivity for the receptor was consistently stronger in myometrium than in leiomyomas during the proliferative phase (Anania et al., unpublished results). In the endometrium, the glandular epithelial cells showed strong cytoplasmic staining during the proliferative and early-mid secretory phase of the cycle, with reduced staining at the time of menstruation. In contrast, stromal cells showed positive staining during the early proliferative phase, but little if any immunoreactivity during the late proliferative and early secretory phase of the cycle. During the mid- to late-secretory phase, the decidualized stromal cells showed dark perinuclear or cytoplasmic staining. Thus, there is differential regulation of the FGF receptor as well. Additionally, there is a secreted form of the FGFR-1 that may act as an bFGF binding protein, and we have identified this form to be present in the leiomyomatous uterus (Hanneken and Baird, 1995; Anania et al., unpublished results).
The type II FGF receptor (FGFR-2) is also a
tyrosine kinase, encoded by the bek gene (Fernig and Gallagher, 1995).
FGFR-2 transduces the actions of both bFGF and acidic FGF. The mRNA for FGFR-2
has been has been identified in both leiomyomas and myometrium (Pekonen et
al., 1993).
Vascular endothelial growth factor
VEGF is an angiogenic growth factor that is a potent mitogen for endothelial cells (Charnock-Jones et al., 1993). It is an approximately 46-kd dimeric protein with four alternatively spliced mRNA species that are all found in human endometrium and myometrium (Charnock-Jones et al., 1993). The receptor for VEGF, Flk-1 (also known as fms-like tyrosine kinase-1) is a 180 kd tyrosine kinase (Seetharam et al., 1995). The expression of both VEGF and Flk-1 correlate temporally and spatially with endothelial cell proliferation in a rat model (Cullinan-Bove and Koos, 1993).
There appears to be menstrual cycle-specific
expression of VEGF in the uterus. During the proliferative phase, VEGF mRNA was
detected in distinct cells in the stroma by in situ hybridization, with weak
expression in the glands (Charnock-Jones et al., 1993). VEGF mRNA was
also detected in the myometrium with the strongest intensity of hybridization at
the endometrial-myometrial border (Charnock-Jones et al., 1993). In
contrast, during the secretory phase there was increasing expression in the
glands, peaking in menstrual endometrium with disappearance of stromal
expression (Charnock-Jones et al., 1993). Evidence from the human fetal
model suggests that VEGF is secreted by epithelial cells and myocytes to
modulate the function of endothelial cells in a paracrine fashion (Shifren et
al., 1994). In the human uterus, VEGF levels were found to be similar in
both myometrium and leiomyomas and to have no significant menstrual cycle
variability (Harrison-Woolrych et al., 1995).
Heparin-binding epidermal growth factor
HBEGF is a 22-kd peptide that functions as a mitogen for both fibroblasts and smooth muscle cells (Higashiyama et al., 1991). HBEGF has an EGF-like domain and thus binds to the EGF receptor (EGF-R) which transduces its action (Higashiyama et al., 1991). In fact, HBEGF has a greater affinity for the EGF-R on smooth muscle cells than does EGF, and thus is a more potent mitogen (Higashiyama et al., 1991). HBEGF expression is elevated at sites of wound healing (Marikovsky et al., 1993) and is a very potent chemoattractant for smooth muscle cells (Higashiyama et al., 1993). In the rat and mouse uterus, HBEGF appears to be modulated by changes in steroid hormone concentrations. Progesterone stimulates HBEGF expression in uterine stromal cells and decreases expression in lumenal and glandular epithelial cells (Zhang et al., 1994; Wang et al. 1994). In contrast, estradiol increases HBEGF expression in epithelial cells and has no effect on stromal cells (Zhang et al., 1994; Wang et al. 1994). Thus, HBEGF appears to be differentially regulated in the endometrium with increased expression correlating with the time of proliferation of a particular uterine cell type. Therefore it may be a mediator of steroid hormone action in the uterus. HBEGF has also been shown to be present in human endometrium, decidua and placenta using reverse transcription polymerase chain reaction (Birdsall et al., 1996).
HBEGF is produced by smooth muscle cells in both leiomyomas and myometrium (Mangrulkar et al., 1995). It is interesting that the levels of mRNA and protein are significantly lower in the neoplastic leiomyomas than in the normal tissue. Immunohistochemistry results show that HBEGF exists primarily as a transmembrane protein in the smooth muscle cells (Ishikawa et al., 1995).
The EGF-R that transduces the actions of HBEGF is
a 170 kd tyrosine kinase receptor (Gill et al., 1985). The C-terminal
region of the EGF-R contains the kinase activity and is homologous to the erb-B
proto-oncogene (Gill et al., 1985). An analysis of EGF-R expression in
human endometrium has shown that epithelial cells express the receptor
throughout the menstrual cycle, while the stromal cells show expression only
during the secretory phase (Ishikawa et al., 1994).
Platelet-derived growth factor
PDGF is a dimeric growth factor with both homodimeric (AA and BB) and heterodimeric (AB) forms linked by disulfide bonds (Dalla Favera et al., 1982; Betsholtz et al., 1986). PDGF has a cysteine-knot structure like that of nerve growth factor, TGF-b and human chorionic gonadotropin (Patel, 1994). Two PDGF receptors have been identified: PDGF-a, which binds all three dimeric hormones, and PDGF-b, which binds only the BB isoform with high affinity (Hart et al., 1988). Both receptors are tyrosine kinases.
Functionally, PDGF acts as a mitogen and chemoattractant for both smooth muscle cells and fibroblasts (Ross et al., 1986). Messenger RNA for both chains has been identified in the human uterus (Boehm et al., 1990; Mendoza et al., 1990), and the protein has been localized to uterine smooth muscle cells by immunohistochemistry (Rossi et al.,1992). The §-isoform of the receptor has also been localized to smooth muscle cells in uterine leiomyomas (Palman et al.,1992).
Studies isolating factors in leiomyomas and
myometrium that are mitogenic for smooth muscle cells and fibroblasts utilizing
the technique of fast protein liquid chromatography (FPLC), have identified PDGF
as a consistent peak (Mangrulakar et al., 1995). Northern blot analysis
of these same tissues showed that both normal myometrial and leiomyoma tissues
expressed the mRNAs for both PDGF chains, with similar levels in both tissues (Mangrulkar
et al., 1995). Immunohistochemistry localized PDGF protein within the
cytoplasm of both fibroid and myometrial smooth muscle cells, and within
vascular smooth muscle cells that line the blood vessels in these tissues. The
intensity of staining was similar for myometrium and leiomyomas.
Transforming growth factor-b
The TGF-b family comprises five dimeric polypeptides encoded by distinct but closely related genes (Barnard et al., 1990; Massague, 1990). TGF-b1,-b2 and -b3 have been identified in a variety of normal and transformed mammalian cells and tissues (Barnard et al., 1990; Massague, 1990; Danielpour et al., 1989). TGF-bs are multifunctional growth factors that regulate many aspects of cellular function including proliferation, differentiation, ECM production, and chemotaxis (Barnard et al., 1990; Massague, 1990; Danielpour et al., 1989). Three distinct proteins have been identified as TGF-§ receptors and have been designated receptor types I-III (Lyons et al., 1991; Cheiftez et al., 1990; Massague, 1992). The type I and II receptors have been shown to be serine/threonine kinases. The type III receptor, also called endoglin, appears to act primarily as a cell surface binding protein (Wrana et al., 1992; Lin et al., 1992). In human endometrium, TGF-§ mRNA is found in both the glandular epithelial and stromal cells, with highest levels present during the late proliferative and early- to mid-secretory phase of the cycle (Tang et al., 1994).
mRNA for TGF-b1 -b2 and -b3 as well as for all three receptors has been detected in both leiomyomas and myometrium (Chegini et al., Murphy et al., 1994). TGF-§1 appears to be a more potent inhibitor of myometrial cell proliferation than of leiomyomas cell proliferation (Murphy et al., 1994). This inhibition may be mediated through the type II receptor since leiomyomas have significantly lower amounts of this receptor than normal myometrial cells (Murphy et al., 1994).
TGF-b3
has been shown to cause increased proliferation of both leiomyoma and myometrial
cells, and a neutralizing antibody to this peptide causes a decrease in
proliferation (Arici et al. 1995, Mayer et al., 1995). Leiomyomas
express up to sixfold higher levels of mRNA for TGF-b3
than do normal myometrial cells at all stages of the menstrual cycle (Murphy et
al., 1994). Thus, TGF-b3
may contribute to the enhanced growth of leiomyomas by increasing cell
proliferation as well as potentially increasing ECM production. Alternatively,
the decreased levels of TGF-b
type II receptor may allow for decreased growth inhibition by TGF-b1.
Parathyroid hormone-related protein
PTHrP is a protein with N-terminal homology to parathyroid hormone (PTH) that was first identified as one of the agents responsible for hypercalcemia of malignancy. The protein and mRNA for PTHrP have subsequently been identified in a variety of normal tissues. In the myometrium of the pregnant rat uterus, PTHrP gene expression is upregulated when the uterus is distended (Daifotis et al., 1992). Data from our laboratory have shown that both PTHrP mRNA and protein are relatively overexpressed in leiomyomas compared with myometrium from the same uterus (Weir et al., 1994). In addition, in both leiomyomas and normal myometrium PTHrP produces a dose-dependent increase in cAMP, which suggests that PTHrP may indeed act as an autocrine regulatory factor for these cells (Weir et al., 1994).
Functionally, PTHrP has been shown to relax smooth muscle and to be a potent vasorelaxant (Daifotis et al., 1992, Botella et al., 1994 ). Vascular smooth muscle cells appear to possess specific receptors for this peptide (Okano et al., 1994). Thus, PTHrP may effect leiomyoma related bleeding by decreasing vascular tone. However, PTHrP has also been reported to have biological effects similar to those of TGF-b, particularly its effects on matrix production (Insogna et al., 1989; Eielson et al., 1994). An alternative mechanism for PTHrP action is that the composition of supporting ECM matrix around the vessels my be changed so that the vascular tone is altered through this indirect action.
Both PTHrP and PTH act via a receptor with
7-transmembrane spanning domains that is capable of stimulating both adenylate
cyclase and phospholipase C (Abou-Samra et al., 1992). This receptor
recognizes the N-terminal region of the ligand (Abou-Samra et al., 1992).
There is also evidence that there may be a second receptor that recognizes
another region of the PTHrP molecule (Rodda et al., 1988).
Prolactin
The human uterus is the major extrapituitary source of prolactin with endometrium, myometrium, leiomyomas and decidua of pregnancy all demonstrating prolactin protein synthesis (Maslar et al., 1980; Walters et al., 1983; Daly et al., 1984). Recent work has established that prolactin functions as an autocrine or paracrine growth factor in myometrium and leiomyomas (Mora et al., 1995). Functionally, prolactin has also been demonstrated to act as a mitogen for uterine smooth muscle cells (Kornyei et al., 1993). A cleaved form of prolactin, the 16-kd fragment, appears to play an important role as an inhibitor of angiogenesis (Folkman, 1996) and acts to inhibit endothelial cell proliferation and chemotaxis (Clapp et al.,1993). Further studies will need to be conducted in uterine tissues to determine whether this active fragment is present and regulates angiogenesis in these tissues.
The prolactin receptor has a single transmembrane
domain and is homologous to the growth hormone receptor (Boutin, et al.,
1988). The postrecptor mechanisms for this receptor are not well understood
(Kelly et al., 1991). We have recently identified the presence of the
prolactin receptor in both leiomyomas and myometrium (Nowak, et al.,unpublished
results).
Conventional therapies for leiomyomas
Current strategies for dealing with leiomyomas and specifically problems with abnormal uterine bleeding that arise from them, fall into two broad categories of therapy. The first is surgical therapy, traditionally hysterectomy or myomectomy but also more recently endoscopic procedures aiming for complete or partial removal of the leiomyomas. While there will always be a role for surgery in some patients, clearly, it is not optimal therapy for all patients.
The second strategy for the management of leiomyomas is manipulation of the steroid hormone environment. The most widely used agent, gonadotropin-releasing hormone agonist (GnRH-a) creates a hypogonadotropic hypogonadal environment that many studies have shown produces amenorrhea and thus controls bleeding (Stewart and Friedman, 1992). In addition GnRH-a is able to induce a significant decrease in uterine size in most women (Stewart and Friedman, 1992). The limitations of GnRH-a therapy for the control of leiomyoma-related bleeding are threefold. First, and most important, this therapy affects all estrogen dependent tissues and thus causes significant hypoestrogenic symptoms; thus long-term therapy is prohibited because significant hypoestrogenic effects on bone can lead to osteoporosis (Stewart and Friedman, 1992). Second, upon discontinuation of therapy, there is rapid regrowth of the uterus often to sizes above pretreatment levels (Stewart and Friedman, 1992). Finally, in a small proportion of women, vaginal hemorrhage can be precipitated by this therapy (Friedman,1989).
Other therapies, including high dose progestin therapy, danazol and gestrinone and oral contraceptives act chiefly on the endometrium to produce atrophy (Rein and Friedman, 1988). These agents tend to be less effective in the control of bleeding than GnRH-a and/or to have significant adverse effects. Therapies directed against the endometrium are supported by data that suggest that in some patients the endometrial lining overlying submucosal fibroids is abnormal (Paterson-Keels et al., 1994).
The possibility of utilizing specific hormone
antagonists, rather than completely shutting down of the
hypothalamic-pituitary-ovarian axis raises the prospect of more selective
therapy. Recent studies suggest that myometrial metabolism is different in
menopausal women than in women treated with GnRH-a; thus other factors,
including gonadotropins may also be regulating tissues traditionally regarded to
be steroid-responsive (Stewart et al, 1995). Pilot studies of the hormone
antagonist mifepristone (RU-486) suggest that it may be a better agent for
leiomyomas than GnRH-a because equivalent reductions in uterine size and
menstrual suppression are obtained without the profound hypoestrogenism
associated with GnRH-a therapy (Murphy et al., 1993 and 1995).
Innovative therapies based on molecular
mechanisms of action
The more clearly the molecular mechanisms for leiomyoma-related menorrhagia are defined, the better therapy can specifically target this problem. As is true for many other clinical disorders, the presence of a specific gene defect would open the possibility of gene therapy in the future. Before gene therapy becomes widely used, however, we will probably pass through an era of therapy directed against specific growth factors involved in the process of angiogenesis within the endometrium and myometrium.
A number of molecules have been identified as inhibiting the process of endothelial cell proliferation and thus inhibiting angiogenesis (Folkman, 1996). Both TGF-b and the secreted form of the bFGF receptor are present in uterus and act as inhibitors of this process (Folkman, 1996). In addition, the 16-kd fragment of prolactin, angiostatin, thrombospondin-1, platelet factor 4, tissue inhibitors of metalloproteinases (TIMPs 1,2,and 3), interferon-a and placental proliferin-related protein negatively regulate angiogenesis and could be exploited therapeutically (Folkman, 1996).
Pharmacologic agents that counteract angiogenic
factors - drugs that block production of these factors, bind or cleave their
active forms, or interfere with binding to their receptor - can also be useful.
Stimulators of angiogenesis that are potential targets for the development of
antagonists include TGF-b, bFGF, VEGF, and PDGF as discussed
previously (Folkman, 1996). In addition other factors including acidic FGF,
angiogenin, TGF-a, tumor necrosis factor-a,
granulocyte colony-stimulating factor, placental growth factor, platelet-derived
endothelial cell growth factor, interleukin-8, hepatocyte growth factor and
proliferin may be useful targets if further studies suggest that they play a
role in uterine angiogenesis (Folkman, 1996, Osuga et al., 1995).
Conclusion
The targeting of angiogenic molecules is clinically desirable since angiogenesis is usually a pathologic process, except in the female reproductive system. Thus, many of the agents might be administered with minimal adverse systemic effects.
Further investigation into this clinically
important and biologically unique process is clearly warranted. By better
understanding the derangements in vascular function in the leiomyomatous uterus,
we can provide better therapy for women suffering from leiomyoma-related
menorrhagia.
Acknowledgements
This work was supported by the Berlex Scholar
Award, The Berlex Foundation, New York, New York (to E.A.S.) and by HD-30496,
National Institutes of Health, Bethesda, MD (to R.A.N.).
References
Abou-Samra, AB, Juppner, H, Force, T, et al. (1992) Expression cloning of a common receptor for parathyroid hormone and parathyroid hormone-related peptide from rat osteoblast-like cells: a single receptor stimulates intracellular accumulations of both cAMP and inositol triphosphates and increases intracellular free calcium. Proc. Nalt. Acad. Sci. USA, 89, 2732-2736.
Anania,C.A., Stewart,E.A. and Quade,B., et al. (1995) Human endometrial, myometrial and leiomyoma cells express an alternatively spliced form of fibroblast growth factor receptor type I. J. Soc. Gynecol. Invest, 2, 415.
Arici,A., Sozen,I. and Olive,D. (1995) Transforming growth factor-b3 (TGF-b3) promotes proliferation in myometrial and leiomyoma cells. J. Soc. Gynecol. Invest., 2, 411.
Barbieri,R.L., Andersen,J. (1992) Uterine leiomyomas: The somatic mutation theory. In: Barbieri RL (ed) Seminars in Reproductive Endocrinology. New York: Thieme Inc., 10, 301-309.
Barnard,J.A., Lyons,R.M. and Moses,H.L. (1990) The cell biology of transforming growth factor b. Biochem. Biophys. Acta., 1032, 79-87.
Betsholtz,C., Johnson,A. and Heldin,C-H., et al. (1986) cDNA sequences and chromosomal localization of human platelet-derived growth factor A-chain and its expression in tumor cell lines. Nature (Lond), 320, 695-699.
Birdsall, MA, Hopkisson, JF, and Grant, KE. (1996) Expression of heparin-binding epidermal growth factor messenger RNA in the human endometrium. Mol. Hum. Reprod., 2, 31-34.
Boehm,K.D., Daimon,M. and Gorodeski,I.G., et al. (1990) Expression of the insulin-like and platelet-derived growth factor genes in human uterine tissues. Mol. Reprod. Dev., 27, 93-101.
Botella,A., Rekik, M. and Delvaux,M., et al. (1994) Parathyroid hormone (PTH) and PTH-related peptide induce relaxation of smooth muscle cells from guinea pig ileum: Interaction with vasoactive intestinal peptide receptors. Endocrinology, 135, 2160-2167.
Boutin, JM, Jolicoeur, C, Okamura, H, et al. (1988) Cloning and expression of the rat prolactin receptor, a member of the growth hormone/prolactin receptor gene family. Cell, 53, 69-77.
Brandon,D.D., Bethea,C.L., Strawn,E.Y., et al. (1993) Progesterone receptor messenger ribonucleic acid and protein are overexpressed in human uterine leiomyomas. Am. J. Obstet. Gynecol., 169, 78-85.
Bulun,S.E., Simpson,E.R. and Word,R.A. (1994) Expression of the CYP19 gene and its product aromatase cytochrome P450 in human uterine leiomyoma tissues and cells in culture. J. Clin. Endocrinol. Metab., 78, 736-743. Buttram Jr.,V.C., Reiter RC. (1981) Uterine leiomyomata: etiology, symptomatology, and management. Fertil. Steril., 36, 433-445.
Chandrasekhar,Y., Heiner, J and Osuamkpe,C., et al. (1992) Insulin-like growth factor I and II binding in human myometrium and leiomyomas. Obstet. Gynecol., 166, 64-69.
Charnock-Jones,D.S., Sharkey,A.M. and Rajput-Williams,J., et al. (1993) Identification and localization of alternatively spliced mRNAs for vascular endothelial growth factor in human uterus and estrogen regulation in endometrial carcinoma cell lines. Biol. Reprod., 48, 1120-1128.
Chegini, N, Zhao, Y, William, SR. et al. (1994) Human uterine tissue throughout the menstrual cycle expresses transforming growth factor-b1 (TGFb1), TGFb2, TGFb3 type II receptor messenger ribonucleic acid and protein and contains [125I]TGFb1-binding sites. Endocrinology, 135, 439-449.
Cheiftez,S., Hernandez,H. and Laiho,M., et al. (1990) Distinct transforming growth factor-B receptor subsets as determinants of cellular responsiveness to three TGF-b isoforms. J. Biol. Chem., 265, 20533-20538.
Clapp,C., Martial,J.A. and Guzman,R.C., et al. (1993) The 16-kilodalton N-terminal fragment of human prolactin is a potent inhibitor of angiogenesis. Endocrinology, 133, 1292-1299.
Cullinan-Bove,K., Koos,R.D. (1993) Vascular endothelial growth factor/vascular permeability factor expression in the rat uterus: rapid stimulation by estrogen correlates with estrogen-induced increases in uterine capillary permeability and growth. Endocrinology, 133, 829-837.
Daifotis,A.G., Weir,E.C. and Dreyer,B.E., et al. (1992) Stretching induced parathyroid hormone-related peptide gene expression in the rat uterus. J. Biol. Chem., 267, 2345-2348.
Dalla Favera,R., Gallo,R.C. and Giallongo A., et al. (1982) Chromosomal localization of the human homolog (c-sis) of the simian sarcoma virus onc gene. Science, 218, 686-688.
Daly,D.C., Walters,C.A. and Prior,J.C., et al. (1986) Prolactin production from proliferative phase leiomyoma. Am. J. Obstet. Gynecol., 148, 1059-1063.
Danielpour,D., Dart,L.L. and Flanders,K.C., et al. (1989) Immunodetection and quantitation of two forms of transforming growth factor-b (TGF-b1 and TGF-b2) secreted by cells in culture. J. Cell. Physiol., 138, 79-86.
Eielson,C., Kaplan,D. and Mitnick,M.A., et al. (1994) Estrogen modulates parathyroid hormone-induced fibronectin production in human and rat osteoblast-like cells. Endocrinology, 135, 1639-1644.
Farrer-Brown G., Beilby,JOW. and Tarbit M.H. (1971) Venous changes in the endometrium of myomatous uteri. Obstet. Gynecol., 38, 743-751.
Faulkner,R.I. (1944) The blood vessels of the myomatous uteri. Am. Obstet. Gynecol., 47, 185-197.
Fernig, DG and Gallagher, JT. (1994) Fibroblast growth factors and their receptors: an information network controlling tissue growth, morphogenesis and repair. Progr. Growth Factor Res., 5, 353-377.
Ferriani,R.A., Charnock-Jones,D.S. and Prentice,A., et al. (1993) Immunohistochemical localization of acidic and basic fibroblast growth factors in normal human endometrium and endometriosis and the detection of their mRNA by polymerase chain reaction. Hum. Reprod., 8, 11-16.
Folkman,J. (1996) Clinical applications of research on angiogenesis. N. Eng. J. Med., 333, 1757-1763.
Friedman,A.J. (1989) Vaginal hemorrhage associated with degenerating submucous leiomyomata during leuprolide acetate treatment. Fertil. Steril., 52, 152-154.
Gill, GN, Kawamoto, T, Weber, W, et al. (1985) Characterization of the epidermal growth factor receptor. In Murakami, H, Yamane, I, Barnes, DW,et al. (eds), Growth and Differentiation of Cells in Defined Environment. Springer-Verlag, New York, NY, USA, pp. 405-410.
Gloudemans,T., Prinsen,I. and Van Unnik,J.A., et al. (1990) Insulin-like growth factor gene expression in human smooth muscle tumors. Cancer Res., 50, 6689-6695.
Guttmacher,A.E., Marchuk,D.A. and White, R.I. (1995) Hereditary hemorrhagic telangiectasia. N. Engl. J. Med, 333, 918-924.
Hanneken A., and Baird A. (1995) Soluble forms of the high-affinity fibroblast growth factor receptor in human vitreous fluid. Invest. Ophthalmol. Vis. Sci., 36, 1192-1196.
Harrison-Woolrych,M.L., Charnock-Jones,D.S. and Smith,S.K. (1994) Quanitification of messenger ribonucleic acid for epidermal growth factor in human myometrium and leiomyomata using reverse transcriptase polymerase chain reaction. J. Clin. Endocrinol. Metab., 78, 1179-1184.
Harrison-Woolrych,M.L., Sharkey,A.M. and Charnock-Jones,D.S., et al. (1995) Localization and quantification of vascular endothelial growth factor messenger ribonucleic acid in human myometrium and leiomyomata. J. Clin. Endocrinol. Metab., 80, 1853-1858.
Hart,C.E., Forstrom,J.W. and Kely,J.D., et al. (1988) Two classes of PDGF receptor recognize different isoforms of PDGF. Science, 240, 1529-1531.
Higashijama,S., Abraham,J.A. and Klagsbrun,M. (1993) Heparin-binding EGF-like growth factor stimulation of smooth muscle cell migration: dependence on interactions with cell surface heparin sulfate. J. Cell Biol., 122, 933-940.
Higashiyama,S., Abraham,J.A. and Miller,J., et al. (1991) A heparin-binding growth factor secreted by macrophage-like cells that is related to EGF. Science, 251, 936-939.
Insogna,K.L., Stewart,A.F. and Morris,C.A., et al. (1989) Native and a synthetic analogue of the malignancy-associated parathyroid hormone-like protein have in vitro transforming growth factor-like properties. J. Clin. Invest., 83, 1057-1060.
Ishikawa,M., Takashima,S. and Stewart,E.A., et al. (1995) Heparin-binding epidermal growth factor and its receptor are differentially expressed in human endometrium, myometrium and leiomyomas. J. Soc. Gynecol Invest., 2, 417.
Kaiserman-Abramof,I.R., Padykula,H.A. (1989) Angiogenesis in the postovulatory primate endometrium: the coiled arteriolar system. Anat. Rec., 224, 479-489.
Kelly, PA, Ali, S, Rozakis, M, et al. (1993) The growth hormone/prolactin receptor family. Rec. Progr. Horm. Res., 48, 123-164.
Kornyei,J.L., Lei,Z.M. and Rao,C.V. (1993) Human myometrial smooth muscle cells are novel targets of direct regulation by human chorionic gonadotropin. Biol. Repod., 49, 1149-1157.
Lin,H.Y., Want, X.F. and Ng-Eaton,E., et al. (1992) Expression cloning of the TGFB type II receptor, a functional transmembrane serine/threonine kinase. Cell, 68, 775-785.
Lindner,V., Reidy,M.A. (1991) Proliferation of smooth muscle cells after vascular injury is inhibited by an antibody against basic fibroblast growth factor. Proc. Natl. Acad. Sci. USA., 88, 3739-3743.
Lyons,R.M., Millert,D.A. and Graycor,J.L., et al. (1991) Differential binding of TGFb1, -b2, -b3 by fibroblasts and epithelial cells measured by affinity cross-linking of cell surface receptors. Mol. Endocrinol., 5, 1887-1896.
Mangrulkar,R.S., Ono, M. and Ishikawa,M., et al. (1995) Isolation and characterization of heparin-binding growth factors in human leiomyomas and normal myometrium. Biol. Reprod., 53, 636-646.
Marikovsky,M., Brewing,K. and Liu,Py., et al. (1993) Appearance of heparin-binding EGF-like growth factor in wound fluid as a response to injury. Proc. Natl. Acad. Sci. USA, 90, 3889-3893.
Maslar,I.A., Kaplan,B.M. and Luciano,A.A., et al. (1980) Prolactin production by the endometrium of early human pregnancy. J. Clin. Endocrinol. Metab., 51, 78-83.
Massague,J. (1990) The transforming growth factor-b family. Ann. Rev. Cell. Biol., 6, 597- 641.
Massague,J. (1992) Receptors for the TGF-b family. Cell, 69, 1067-1070.
Mayer,J.C., Murphy,J.M. and Tsibris,J.C.M., et al. (1995) Growth of human uterine leiomyoma cells in response to transforming growth factor-b3 varies depending on in vivo estrogen status. J. Soc. Gynecol. Invest., 2, 416.
McAllister,K.A., Grogg,K.M. and Johnson,D.W., et al. (1994) Endoglin, a TGF-b binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type I. Nat. Genet., 8, 345-351.
Mendoza,A.E., Young,R. and Orkin,S.H., et al. (1990) Increased platelet-derived growth factor. A-chain expression in human uterine smooth muscle cells during the physiologic hypertrophy of pregnancy. Proc. Natl. Acad. Sci. USA., 87, 2177-2181.
Mora,S., Diehl,T. and Stewart,E.A., et al. (1995) Prolactin is an autocrine growth regulator for human myometrial and leiomyoma cells. J. Soc. Gynecol. Invest., 2, 396.
Murphy,A.A., Kettel,L.M. and Morales,A.J., et al. (1993) Regression of uterine leiomyomata in response to the anti-progesterone RU486. J. Clin. Endocrinol. Metab., 76, 513-517.
Murphy,A.A., Morales,A.J. and Kettel,L.M., et al. (1995) Regression of uterine leiomyomata to the antiprogesterone RU486: dose response effect. Fertil. Steril., 64, 187-190.
Murphy,J., Tsibris, J. and Tsibris. A., et. al. (1994) Regulation by estrogen: the transforming growth factor b system in uterine leiomyomas. Soc. Gynecol. Invest.. Ann. Meeting, Chicago, May, Abstract P21.
Noyes,R.W., Hertig,A.T. and Rock,J. (1950) Dating the endometrial biopsy. Fertil.Steril., 1, 3-25.
Okano,K., Wu,S. and Huang,X., et al. (1994) Parathyroid hormone (PTH)/PTH-related protein (PTHrP) receptor and its messenger ribonucleic acid in rat aortic vascular smooth muscle cells and UMR osteoblast-like cells: cell-specific regulation by angiotensin-II and PTHrP. Endocrinology, 135, 1093-1099.
Osugo, Y, Toyashima, H, Mitsuhashi, N, et al. (1995) The presence of platelet-derived endothelial cell growth factor in human endometrium and its characteristic expression during the menstrual cycle and early gestational period. Hum. Reprod., 10, 989-993.
Palman,C., Bowen-Pope,D.F. and Brooks,J.J. (1992) Platelet-derived growth factor receptor (b-subunit) immunoreactivity in soft tissue tumors. Lab. Invest., 66, 108-115.
Patel,D.J. (1994) Glycoprotein hormones. A clasped embrace. Nature, 369, 438-439.
Patterson-Keels,L.M., Selvaggi,S.M. and Haefner,H.K., et al. (1994) Morphologic assessment of endometrium overlying submucosal leiomyomas. J. Reprod. Med., 39, 579-584.
Pekonen, F, Nyman, T, and Rutanen, EM (1993) Differential expression of keratinocyte growth factor and its receptor in the human uterus. Mol. Cell. Endocrinol., 95, 43-49.
Presta,M. (1988) Sex hormones modulate the synthesis of basic fibroblast growth factor in human endometrial adenocarcinoma cells: implications for the neovascularization of normal and neoplastic endometrium. J. Cell. Physiol., 137, 593-597.
Rein,M.S., Friedman,A.J. (1988) Medical therapy for uterine leiomyomata. In: ReproductiveEndocrine Therapeutics, Barbieri RL, Schiff I (Eds). Alan R. Liss, Inc, New York.
Rein,M.S., Friedman,A.J. and Barbieri RL, et al. (1991) Cytogenetic abnormalities in uterine leiomyomata. Obstet. Gynecol., 77, 923-926.
Rein,M.S., Friedman,A.J. and Stuart,J.M., et al. (1990) Fibroid and myometrial steroid receptors in women treated with the gonadotropin-releasing hormone agonist leuprolide acetate. Fertil. Steril., 53, 1018-1023.
Rodda, CP, Kubota, M, Heath, JA, et al. (1988) Evidence for a novel parathyroid hormone-related protein in fetal lamb parathyroid glands and sheep placenta: comparisons with a similar protein implicated in humoral hypercalcemia of malignancy. J. Endocrinol., 117, 261-271.
Ross,R., Raines,E.W. and Bowen-Pope,D.F. (1986) The biology of platelet-derived growth factor. Cell, 46, 155-169.
Rossi,M.J., Chergini,N. and Masterson,BJ. (1992) Presence of epidermal growth factor, platelet-derived growth factor, and their receptors in human myometrial tissue and smooth muscle cells: their action in smooth muscle cells in vitro. Endocrinology, 130, 1716-1727.
Sampson,J.A. The blood supply of uterine myomata. (1912) Surg. Gynecol. Obstet., 14, 215-230.
Seetharam, L, Gotoh, N, Maru Y, et al. (1995) A unique signal transduction from FLT tyrosine kinase, a receptor for vascular endothelial growth factor VEGF. Oncogene, 10, 135-147.
Shifren,J.L., Doldi,N. and Ferrara,N., et al. (1994) In the human fetus, vascular endothelial growth factor is expressed in epithelial cells and myocytes, but not vascular endothelium: implications for mode of action. J. Clin. Endocrinol. Metab., 79, 316-322.
Soules,M.R., McCarty,K.S. Jr. (1982) Leiomyomas: steroid receptor content, variations within normal menstrual cycles. Am. J. Obstet. Gynecol., 43, 6-11.
Stewart,E.A., Friedman,A.J. (1992) Steroidal treatment of myomas: Preoperative and Longterm Medical Therapy. In: Barbieri RL (ed) Seminars in Reproductive Endocrinology. New York: Thieme Inc., 10, 344-357.
Stewart,E.A., Friedman,A.J. and Peck,K., et al. (1994) Relative overexpression of collagen Type I and collagen Type III messenger RNAs by uterine leiomyomas during the proliferative phase of the menstrual cycle. J. Clin. Endocrinol. Metab., 79, 900-908.
Stewart,E.A., Jain,P. and Penglase,M.D., et al. (1995) The myometrium of postmenopausal women produces prolactin in response to human chorionic gonadotropin and a-subunit in vitro. Fertil. Steril., 64, 972-976.
Tabibzadeh, S. (1996) The signals and molecular pathways involved in human menstruation, a unique process of tissue destruction and remodelling. Mol. Hum. Reprod., 2, 77-92.
Tabibzadeh, S, Babaknia, A, Kong, QF, et al. (1995) Menstruation is associated with disordered expression of desmoplakin I/II and cadherin/catenins and conversion of F- to G-actin in endometrial epithelium. Hum. Reprod., 10, 776-784.
Tang,X-M., Zhao,Y. and Rossi,M.J., et al. (1994) Expression of transforming growth factor-b (TGFb) isoforms and TGFb type II receptor messenger ribonucleic acids and proteins, presence of [125I] TGFb1- binding sites, and the effect of TGFbs on endometrial stromal cell growth and protein degradation in vitro. Endocrinology, 135, 450-459.
Tommola, R, Pekonen, F and Rutanen, EM. (1989) Binding of epidermal growth factor and insulin-like growth factor I in human myometrium and leiomyomata. Obstet. Gynecol., 74, 658-662.
Torry,R.J., Rongish,B.J. (1992) Angiogenesis in the uterus: potential regulation and relation to tumor angiogenesis. Am. J. Reprod. Immunol., 27, 171-179.
Vladavsky,I., Folkman,J. and Sullivan,R., et al. (1987) Endothelial cell-derived basic fibroblast growth factor: synthesis and deposition into subendothelial extracellular matrix. Proc. Natl. Acad. Sci. USA., 84, 2292-2296.
Vollenhoven,B.J., Herington,A.C. and Healy,D.L. (1993) Messenger ribonucleic acid expression of the insulin-like growth factors and their binding proteins in uterine fibroids and myometrium. J. Clin. Endocrinol. Metab., 75, 1106-1110.
Walters,C.A., Daly,D.C. and Chapitis,J., et al. (1983) Human myometrium: a new potential source of prolactin. Am. J. Obstet. Gynecol., 147, 639-644.
Wang,X-N., Das,S.K. and Damm,D., et al. (1994) Differential regulation of heparin-binding epidermal growth factor-like growth factor in the adult ovariectomized mouse uterus by progesterone and estrogen. Endocrinology, 135, 1264-1271.
Weir,E.C., Goad,D.L. and Daifotis,A.G., et al. (1994) Relative overexpression of the parathyroid hormone-related protein gene in human leiomyomas. J. Clin. Endocrinol. Metab., 78, 784-789.
Wilcox,L.S., Koonin,L.M. and Pokras, R., et al. (1994) Hysterectomy in the United States, 1988-1990. Obstet. Gynecol., 83, 549-555.
Wrana,J.L., Attisano,L. and Carcamo,J., et al. (1992) TGF b signals through a heteromeric protein kinase receptor complex. Cell, 71, 1003-1014.
Yeh,J., Osathanondh,R. (1993) Expression of messenger ribonucleic acid encoding for basic fibroblast growth factor (FGF) and alternatively spliced FGF receptor in human fetal ovary and uterus. J. Clin. Endocrinol. Metab., 77, 1367-1371.
Zhang,A., Funk, C. and Roy,D., et al. (1994) Heparin-binding epidermal growth factor-like growth factor is differentially regulated by progesterone and estradiol in rat uterine epithelial and stromal cells. Endocrinology, 134, 1089-1094.